
Если следовать аксиомам популяционной генетики, стоит отметить, что на Земле нет чистых популяций (народов). Все народы на Земле родственны друг другу, в той или иной степени. Человечество генетически едино и на 99,99 % генетически идентично, несмотря на расовые и антропологические внешние различия. Все наше различие равняется 0,01 %. Поэтому, не стоит уделять этой маленькой цифре такое большое внимание, как это делают идеологи этнонационализма и ксенофобии.
Генетически казахи тесно связаны со всеми народами Евразии. Это вызвано с одной стороны, тем, что Великая степь была главным коридором масштабных миграций из Азии в Европу и обратно.
А с другой стороны, все проживавшие здесь народы и культуры оставили свой генетический след в общем этногенезе казахов.
Ниже будет прослежена генетическая история Казахстана, основанная на изучении методами популяционной генетики останков представителей разных культур и народов, проживавших на территории Великой степи.
1. Население Великой Степи с точки зрения полиморфизма Y-хромосомы.
Первые результаты палеогенетических исследований полиморфизма Y-хромосомы стали выходить в начале 2000-х годов. Сразу стоит оговорится, что пока данных палеогенетики очень мало. И абсолютизировать данные результаты пока не стоит, так как данная выборка еще малорепрезентативна. Для нормального описания любой древней популяции нужно как минимум 30 образцов (в идеале 1500) принадлежащих к одной эпохе и взятых в разных географических местах. Сейчас, за все время, начиная с начала 2000-х годов, протестировано менее 100 древних образцов за все периоды истории. Причем большая их часть была опубликована в 2015 году. Судя по таким темпам и научным отчетам палеогенетиков, в ближайшее время будут публиковаться все больше новых палеогенетических данных.
К примеру, в лаборатории древнего ДНК в Копенгагене на данный момент исследуются около 1000 древних образцов, из них 150 из Центральной Азии . Палеогенетические исследования стали обретать свою научную значимость и актуальность по мере накопления научных знаний и совершенствования технологий секвенирования древнего ДНК.
Археологические культуры степной части Евразии бронзового века.
Одной из первых археологических культур, которая была изучена популяционными генетиками, была Андроновская археологическая культура, охватывавшая в XVII—IX веках до н. э. Казахстан, Западную Сибирь, западную часть Средней Азии и Южный Урал.
На данный момент выделяют следующие локальные варианты Андроновской культуры:
1. Синташта-Аркаим (Южный Урал, Северный Казахстан, 2200—1600 гг. до н. э.,
1.1. Поселение Синташта, датируемое 1800 г. до н. э.;
1.2. Поселение Аркаим, датируемое 1700 г. до н. э.;
2. Алакуль (2100—1400 гг. до н. э.), в районе между реками Амударья и Сырдарья, пустыня Кызылкум;
2.1. Алексеевка (1300—1100 гг. до н. э.) в восточном Казахстане;
2.2. Ингальская долина на юге Тюменской области, в которой последовательно сменяют друг друга памятники алакульской, федоровской и саргатской культур.
3. Федорово (1500—1300 гг. до н. э.) в Южной Сибири (впервые встречается кремация и культ огня);
4. Бешкентский район — Вахш (Таджикистан), 1000—800 гг. до н. э. На данный момент представители данной археологической культуры наиболее сильно изучены с точки зрения популяционной генетики.
Андроновская археологическая культура.
Одними из первых в мире, стали публиковаться палеогенетические данные по Андроновской археологической культуре. Это связано с ключевым местом данной археологической культуры в научных дебатах по поводу распространения индоевропейских и индоиранских языков.
В своей пионерской работе 2009 года Кайзер и соавторы опубликовали первые палеогенетические данные по представителям Курганной археологической культуры. Ими были взяты образцы 32 представителей Курганной археологической культуры на территории Красноярского края.
Курганная культура - это принятое на Западе общее названия для ряда родственных археологических культур, имеющих примерно одинаковую антропологию и материальную культуру. Из этих 26 останков 10 относились к Андроновской археологической культуре, 4 к Карасукской археологической культуре, 12 к Тагарской археологической культуре, 6 к Таштыкской археологической культуре. Последние три археологические культуры происходят от Андроновской, но мало знакомы неспециалистам из Казахстана, в силу своей географической расположенности не на территории Казахстана возле ее границ. Карасукская культура преемница Андроновской культуры, ей наследовала Тагарская культура, после этого на месте Тагарской культуры возникла Таштыкская культура. Образцы Андроновской культуры были взяты из таких поселений как Татарка, Солеозерная I, Солеозерная IV, Усть-Абаканский. Датировка останков 1800-1400 года до нашей эры. Среди представителей андроновской культуры было получено всего 3 результата. Из 3 протестированных 2 относятся к гаплогруппе R1a, которая имеет западноевразийское происхождение и 1 относится к гаплогруппе С, имеющий восточноазиатское происхождение. Эти данные частично подтверждают данные лингвистов и антропологов об индоевропейском (а точнее индоиранском) происхождении представителей андроновской культуры. Также эти данные позволяют проследить генетические связи с населением древнего и средневекового Казахстана. Гаплогруппа R1a доминирует у многих тюркских народов Евразии (у кыргызов и алтайцев составляет не менее 50 % всей популяции), среди казахов не более 10 %. После этого 6 лет не было никаких новых данных по представителям Андроновской Археологической культуры до публикаций 2015 года.
В статье Кайзер 2009 года присутствуют данные по палеоДНК культур, которые наследовали Андроновской культуре: К сожалению, данные полиморфизма Y-хромосомы по Карасукской культуре не были получены авторами исследования в силу большой деградации ядерного ДНК, но палеогенетические данные по другим археологическим культурам были получены и опубликованы в статье. Образцы Тагарской археологической культуры были взяты на поселениях Анач, Черногорский и Усть-Абаканский, Бейский и Богратский и датируются периодом 800 г. до н.э.- 100 г. н.э. Все 6 останков принадлежат к гаплогруппе R1a. Единственный результат Таштыкской археологической культуры (поселение Абакано-Перевоз I (I-V века)), относится также к гаплогруппе R1a.

Также интересным являются факты, опубликованные в статье по поводу генетического определения цвета глаз у останков людей. Как известно цвет глаз определяется мутацией в ДНК-сегменте rs12913832 . Если в данной позиции стоит Гуанин, то у человека будут голубые глаза, что является поздней мутацией, по сравнению с изначальным Тимином на данной позиции, определяющим серый цвет глаз. Исследовав гены представителей Курганных культур, авторы нашли то, что у 15 человек были голубые глаза, а у 8 серые глаза. Данные генетические открытия коррелируют с данными палеоантропологии, согласно которой, представители данных культур относились к европеоидной расе, с небольшими примесями монголоидной расы.
Таким образом, несмотря на минимальные данные по палеоДНК представителей мы видим четкую генетическую преемственность начиная от периода 1800 года до нашей эры, заканчивая ранним средневековьем.
К сожалению, глубина теста (выявлен общий SNP-маркер гаплогруппы R1a, но не известен субклад) не позволяет уточнить глубину родства с современными популяциями. Идентификация субкладов среди представителей различных археологических культур в научных статьях появилась только недавно, в 2014-2015 годах. Примером более глубокого исследования является нижеследующая научная работа 2015 года. В публикации Аллентофта в июне 2015 года были обозначены следующие данные по представителям Андроновской Археологической культуры: 2 образца из Синташты (2100-1800 лет до н.э.) относятся к гаплогруппе R1a, 4 образца Карасукской культуры (1400-900 лет до н.э.) относятся к гаплогруппам R1a (3 образца) и Q1a (1 образец), 3 образца Андроновской культуры относятся к гаплогруппе R1a, из 5 образцов ямной культуры, 4 принадлежат к гаплогруппе R1b, а еще один к гаплогруппе I2a, 2 представителя Межовской культуры (останки из Каповой пещеры. Современная Башкирия) относятся к гаплогруппам R1a и R1b, из 3 образцов с Алтая (900/700 лет до н.э.-500/1000 г. н.э.) 1 относится к гаплогруппе J2a, а два образца к гаплогруппе Q1a.
Стоит отметить, что, начиная с 2009 года (первая публикация по этим древним популяциям) технологии секвенирования кардинально улучшились. Здесь результаты даны в общем виде, без уточнения субкладов. Биоинформатик Владимир Таганкин проанализировал исходные BAM-файлы и дополнительно предоставил данную информацию: Образец RISE386 (Буланово, Синташта) «датирован 2298-2045 годами н.э., относится к субкладу Z2121/S3410+ Z2124+, YP1460+. Вероятно, образец может относится к субкладу YP1460, параллельному к Z2123. YP1460 является позитивным для киргизской ветви».
Здесь стоит уточнить, что данный субклад предковый для 40 % современных кыргызов. Также сюда относятся некоторые польско-литовские татары. Кроме того, тестированный на полный сиквенс Y-хромосомы казах из рода Табын совпадает с данным субкладом (YF03517). Малое количество тестированных человек на полный сиквенс Y-хромосомы не позволяет сказать, сколько процентов каких популяций совпадает с этим образцом. Предварительные результаты показывают три популяции (кыргызы, казахи, польско-литовские татары), имеющие связи с этим палеогенетическим образцом из Синташты (ранний этап Андроновской культуры). RISE392 (Степное 7, Синташта). «датирован 2126-1896 годами до н.э. Здесь качество сиквенса получше. Образец относится к ветви Z93>Z94>Z2124>Z2125>Z2123>Y877. Он имеет положительный результат по снипу Y877, который расположен на уровне R-Y875. Проверенные снипы этого уровня: Y939- Y877+. SNP Y877 обнаружен в двух научных образцах из популяции Телугу (Индия)».
Здесь стоит отметить, что наиболее близкими данному субкладу являются популяции пуштунов, индусов, башкирских Кыпчаков и Еланов, Карачаево-балкарцев, башкирских Табынов.

Чуть дальше от него расположен казах, отмеченный выше, относящийся к параллельному субкладу. То есть данный образец не предковый, но родственный казахскому образцу YF03517 RISE512 (Кытматово, Андроновская культура), «датирован 1446-1298 годами до н.э. Все снипы ниже Z2124, охваченные севенирванием, отрицательные. Y877 тоже в минусе. R-Z2124: Z2121/S3410+».
Судя по всему, это тупиковая ветвь, не оставившая потомства. Родственными для этого образца являются многие современные тюркские и южноазиатские популяции, но, ни одна из них, не имеет прямого родства с данным образцом. RISE495 (Арбан 1, Карасукская культура), «без датировки. Принадлежит субкладу R-S23592 (Z2124+ Z2125+ Z2122- Z2123-). Определен положительный YP349. Субклад является родительским к киргизской ветви». Здесь можно отметить, что казах YF3517 близко родственен данному образцу, но не является его прямым потомком, в отличии от кыргызов, которые наиболее близки данному образцу. RISE494 (Сабинка 2, Карасукская культура), «датирован 1416-1268 годами до н.э. Сиквенс имеет очень малый охват с низким покрытием. Точно положительный для Z645. Положительное значение для YP521 конфликтует с отрицательным для Z2123. Если YP521+ правильный, то образец может быть отнесен к субкладу Z645>Z93>Z94>Z2124>Z2125>Z2123>Y934> YP521. В настоящее время в этот субклад входит группа индусов и саудитов». Здесь стоит отметить, что данная ветвь могла быть, как и предковой для индусов и арабов (близкие к этому субкладу также башкирские еланы и табыны, а также карачаевцы), так и тупиковой, в которой независимо произошла мутация YP521+. Не очень хорошее качество секвенирования не позволяет точно сказать к какому субкладу относился данный образец. RISE492 Сабинка 2, «датирован 396-209 годами до н.э. M417+ имеет три противоречащих друг другу положительных SNP. R-YP1506 - ветвь объединяет пенджабца, алтайцев и киргизов из Таджикистана. Ветка параллельная к Z94. R-Y46 – индийцы. R-YP294 - Тоскания и Турция». Возможны все три варианты. В первом варианте стоит отметить, что под кыргызами имеют ввиду кыргызов из подразделения Ичкилик. Потенциально к этой ветке близок казахский род Кердери, а также хакасы. Но данная гипотеза требует верификации путем проведения полного сиквенса Y-хромосомы у представителей данных популяций. RISE525 (Капова пещера, Межовская культура), «образец без датировки. Качество плохое и результаты весьма противоречивые. Может даже оказаться Z280 (Y11175+ противоречит Z283-). Ниже отчет по всем покрытым снипа R1a: R-YP541: YP546+ и R-Y11162: Y11175+». Здесь образец в любом из двух случаев явно тяготеет к западноевропейским или восточноевропейским R1a (а не азиатским R1a как все предыдущие). В конце стоит обозначить, что близость к индусам и арабам, отнюдь не означает, что только те популяции близки останкам. Это лишь констатирует тот факт, что на данный момент арабы и индусы более глубоко исследованы, чем тюркские и другие азиатские (в том числе казахская) популяции. Полные сиквенсы 7 палеогенетических останков Андроновской и Карасукской культур показывают прямое генетическое родство казахов рода Табын с образцом из захоронения Буланово (Синташта, Андроновская культура), близкородственные генетические связи с представителем Карасукской культуры и среднеродственные генетические связи с 4 образцами из Андроновской и Карасукской культур. Генетически относительно дальним оказались останки человека из Каповой пещеры, которые больше тяготятся к славянским популяциям. Их общий предок с современными казахами гаплогруппы R1a жил 5500-8000 лет назад.
Новые данные из препринта Яна Матиесона и соавторов, вышедшего 10 октября, показывают немногочисленные результаты по Андроновской археологической культуре: Также были опубликованы исследованные ранее 2 образца (R1a-L62, R1a1a1b-S224) из Синташты (2298-1896 гг. до н,э.) и один образец (R1a1a1b-S224) Андроновской культуры (1446-1298 гг. до н.э.) Идентификация родства осложняется не полной исследованностью современной популяции казахов. На данный момент среди современной казахской популяции исследован всего один субклад гаплогруппы R1a (Казах рода Табын), хотя данных субкладов у казахов насчитывается не меньше 10.
Дальнейшее исследование древнего ДНК Курганных археологических культур и исследование современного населения Казахстана покажет гораздо более плотные и сложные связи между современным населением Казахстана и его древними насельниками. Андроновская археологическая культура доминировала на территории Казахстана, но помимо нее было еще несколько археологических культур, которые захватывали часть территории современного Казахстана и были генетически родственны части современного населения Казахстана. Одной из таких культур была Ямная археологическая культура.
2. Самарская, Ямная и Полтавская археологические культуры.
Самарская археологическая культура начала 5-го тысячелетия до н. э. (эпоха Энеолита) существовала в районе Самарской Луки реки Волга. Обнаружена в 1973 году во время археологических раскопок возле поселения Съезжее. Восточной границей этой культуры был бассейн реки Урал (современная Западно-Казахстанская и Атырауская области). Наследником Самарской археологической культуры называют Ямную археологическую культуру.
Ямная археологическая культура датируется эпохой позднего медного века — раннего бронзового века (3600-2300 гг. до н.э.). Занимала территорию от Южного Приуралья (Западный Казахстан) на востоке до Днестра на западе.
Полтавкинская археологическая культура возникла в начале бронзового века. Датируется 2700-2100 гг. до н.э. Возникла на месте Ямной археологической культуры. Многие исследователи считают ее продолжением Ямной археологической культуры. Генетические результаты представителей данной культуры подтверждают тезис о преемственности от Ямной археологической культуры, добавляя к этому факты миграций с востока, которые фиксируются генетически. Единственное отличие от Ямной культуры заключается в небольшой примеси со стороны Срубной или Андроновской археологических культур.
В июне 2015 года были опубликованы результаты исследований 7 представителей Ямной культуры. Все 7 принадлежат к гаплогруппе R1b1a, из которых один относится к субкладу R1b1a (видимо P297+), один к субкладу R1b1a2а (видимо L23+), еще 5 к субкладу R1b1a2a2 (видимо Z2103+) . К последнему субкладу близки современная популяция казахских баганалы-найманов, проживающих в Центральном Казахстане в районе Улытау.
Новые данные из препринта Яна Матиесона и соавторов, вышедшего 10 октября, дают следующие результаты: Палеогенетические останки из поселения Хвалынск 2: 3 образца Самарской культуры (5200-4000 лет до н.э.) относятся к гаплогруппам R1b1-M415, R1a1-M459, Q1a-F2676. Пять образцов Полтавкинской археологической культуры (датировки укладываются в период 2925-2200 лет до н.э.) относятся к субкладам: R1b1a2a2-Z2105, R1b1a2-L773, R1b1a2a-L23, R1b1a2a2-Z2105, R1a1a1b2a-Z94. Двенадцать образцов Ямной культуры (3330-2143 лет до н.э.) (5 из них публиковались ранее): R1b1a2-PF6482, R1b1a2-L482, R1b1a2a2-Z2105 (4 образца), R1b1a2-PF6399, I2a2a1b1b-L699, R1b1a2a2-CTS1078, R1b1a2-PF6434, R1b1a2a-L23, R1b1a2-PF6409.
Как мы видим 11 из 12 относятся к гаплогруппе R1b1a2-M269. Два образца (2469-1900 лет до н.э.) из поселения Утевка 6 (Самарская область), Полтавкинской археологической культуры относятся к данным гаплогруппам P1-P284 (экзотический вариант), R1a1a1b-S441 (азиатский субклад R1a).
Также была опубликована диссертация, защищенная во Франции в 2014 году, согласно которой представители Афанасьевской культуры в Сибири относятся к той же гаплогруппе R1b1a2, что и Ямная культура. К сожалению субклад не известен, но уже этот факт позволяет предположить, что Ямная и Афанасьевская культуры были родственны.
3. Срубная археологическая культура.
Срубная археологическая культура возникла в эпоху поздней бронзы (XIX-XII века до нашей эры. Основной ареал распространения в степной и лесостепной полосе Степной зоны Евразии между Днепром и Уралом (Западный Казахстан), с отдельными памятниками в Западной Сибири и северном Казахстане.
Данные из препринта Яна Матиесона и соавторов дают следующие результаты по срубной археологической культуре, которая локализуется на границе России и Казахстана: шесть образцов Срубной культуры (1850-1200 лет до н.э.) относятся к следующим субкладам: R1a1a1b2-Z93 (3 образца), R1a1-M459, R1a1a-L168, R1a1a1b2-Z2123 .
Этот факт показывает, что несмотря на географию проживания на месте ямной культуры, генетические связи представителей срубной культуры с ямной археологической культурой не такие большие. В то же время генетические связи срубной археологической культуры сближают ее с Андроновской Археологической культурой.
4. Безымянные Археологические культуры Алтая.
Группа исследователей изучила 14 образцов палеоДНК с Алтайских гор. 13 из них относились к эпохе Бронзового века, а еще один из Энеолитического периода. Захоронения относятся к трем гаплогруппам: Q1a2a1-L54, R1a1a1b2-Z93 and C-М130.
В целом можно отметить, что эти три гаплогруппы в общем среди казахов составляют две трети популяции. Отсутствие субкладов в данном исследовании не позволяет сказать что-либо насчет глубины родства, но сам факт наличия родства фиксируется.
5. Кочевники с территории восточной части Великой степи.
Интересными являются ранние захоронения эпохи династии Чжоу в Китае (XI век до н.э.). Захоронения в Западном Китае (Чэнбэй, столица провинции Цинхай (Кукунор)) относятся к этому периоду. Были исследованы 27 образцов из Чэнбэя эпохи династий Шан и Чжоу (примерная дата захоронений 1000 лет до н.э.). Были выделены 3 социальные группы и получены образцы 27 человек.
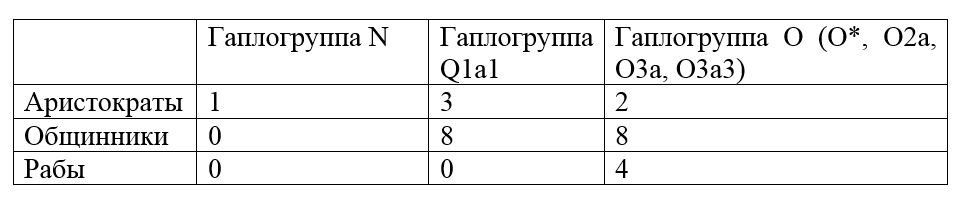
Здесь видно большой процент относящихся к гаплогруппе Q1a1 среди высоких социальных групп. Данная гаплогруппа близка части современных казахов. В то же время исконные китайские субклады (гаплогруппы О), относящиеся к этой эпохе, тяготеют к менее престижным социальным группам. С учетом эпохи Чжоу, географии захоронений (Кукунор), и происхождения гаплогруппы Q1a1 (не автохтоны Китая, а мигранты с Запада, Севера и Северо-Запада), мы можем предположить, что гаплогруппа Q1a1-М120 маркировала чжоуское население, которое не было близко автохтонному китайскому населению, а больше тяготело по происхождению к степным районам Евразии и горного Алтая. Как известно, данная династия была иноземной и смогла завоевать Китай в XI веке до н.э.
Среди отличительных особенностей воинов династии Чжоу было использование колесниц. Этот факт и данные генетических анализов позволяют утверждать, что династия Чжоу была «западной по происхождению» и происходила с территории «Великой степи».
Гаплогруппа Q, найденная у представителей аристократии времен династии Чжоу имеет Алтайско-Сибирское происхождение. Среди казахов, к этой группе относится род Канлы, имеющий древнюю историю на территории Казахстана.

6. Ботайская культура.
Один образец Ботайской культуры из Северного Казахстана (недалеко от с. Никольское, р. Иман-Бурлук) датирован IV-III тысячелетием до н.э. Череп из группового захоронения (раскоп XIV, 1983 г.) был секвенирован в лаборатории Общей генетики и цитологии (Алматы), где был установлен 17-маркерный STR-гаплотип: DYS390-24, DYS391-11, DYS392-13, DYS393 - 14, DYS19 - 15, DYS385 a/b - 17/18, DYS439 - 13, DYS389 I - 12, DYS389 II - 29, DYS448-23, DYS458 - 15, DYS437 - 15, GATA H4 - 11, DYS456 - 16, DYS438 - 13, DYS635 – 21. К сожалению, гаплогруппа не была установлена. Предикторное значение гаплогруппа О2 ошибочно в силу несовершенства самого предиктора.
Судя по разным версиям экспертов данный гаплотип относится к гаплогруппе Q, о которой мы писали выше. Также данной лабораторией была установлена женская гаплогруппа - K1b2 гаплотип мтДНК .
Помимо этого, стоит упомянуть также о генетических связях народов Великой степи с населением Северной и Южной Америк. Большая часть автохтонного населения Северной и Южной Америк относится к гаплогруппе Q, а точнее к ее субкладу Q1a2a1a1-M3. Данный субклад еще 13100 лет назад пересек Берингов пролив и начал заселять Америку. Но помимо этой миграции из Сибири, была еще вторая миграция, более поздняя по времени. Потомки второй волны миграции из Азии в Северную Америку относятся к субклады С2-P39 . Данная миграция не была такой глобальной как миграция гаплогруппы Q, она лишь затронула часть Северной Америки. К данному субкладу относятся американские индейцы Атабаскской языковой семьи. Они населяют западные районы США и Канады. Самыми известными племенами из этой группы были Апачи и Навахо.
Здесь стоит отметить, что данный субклад С2-Р39 довольно близок палеогенетическим останкам из прототюркских (хуннских погребений), причем ближе чем субклады гаплогруппы С2 современных монголов, корейцев и северных китайцев.
Таким образом, мы видим генетические связи между кочевыми народами Великой степи и другими народами Евразии и Нового света.
Подводя итоги можно сказать, что мы пока находимся на стадии накопления научных палеогенетических данных о древнем населении степной части Евразии. Выборки на данный момент мало репрезентативны с количественной точки зрения, но даже они сейчас помогают приоткрыть завесу тайны над генетическим происхождением древнего населения Великой степи, а также проследить генетические связи между древним и современным населением Казахстана.
Жаксылык САБИТОВ,
доктор PhD
Источники:
Johannes Krause, Ludovic Orlando, David Serre, Bence Viola, Kay Prüfer, Michael P. Richards, Jean-Jacques Hublin, Catherine Hänni, Anatoly P. Derevianko, Svante Pääbo. Neanderthals in Central Asia and Siberia//Nature. 449. 2007, pp. 902—904
The Neandertal Adolescent Le Moustier 1. New Aspects, New Results// H.Ullrich (Ed.). Berliner Beitrage zur Vor – und Fruhgeschichte, N.F., Bd.12. Berlin, 2005, 354 p. 5TH ANNUAL MEETING OF THE European Society for the study of Human Evolution. 10-12 September. 2015. http://www.eshe.eu/static/eshe/files/PESHE/PESHE_4_2015_London.pdf
Richard Green et al. A Draft Sequence of the Neandertal Genome//Science. 2010. 07 may (vol. 328, no. 5979). P. 710–722.
M. Gallego Llorente, E. R. Jones, A. Eriksson, V. Siska, K. W. Arthur, J. W. Arthur, M. C. Curtis, J. T. Stock, M. Coltorti, P. Pieruccini, S. Stretton, F. Brock, T. Higham, Y. Park, M. Hofreiter, D. G. Bradley, J. Bhak, R. Pinhasi, A. Manica. Ancient Ethiopian genome reveals extensive Eurasian admixture throughout the African continent//Science. Published Online October 8 2015. Richard Green et al. A Draft Sequence of the Neandertal Genome//Science. 2010. 07 may (vol. 328, no. 5979). P. 710–722.
Condemi S, Mounier A, Giunti P, Lari M, Caramelli D, Longo L. Possible interbreeding in late Italian Neanderthals? New data from the Mezzena jaw (Monti Lessini, Verona, Italy)//PLoS One. 2013; 8(3): e59781. Epub 2013 March 27. http://www.ncbi.nlm.nih.gov/pubmed/23544098
Johannes Krause, Qiaomei Fu, Jeffrey M. Good, Bence Viola, Michael V. Shunkov, Anatoli P. Derevianko & Svante Pääbo The complete mitochondrial DNA genome of an unknown hominin from southern Siberia//Nature 464, 894-897 (8 April 2010). Published online 24 March 2010; http://www.nature.com/nature/journal/v464/n7290/abs/nature08976.html 5TH ANNUAL MEETING OF THE European Society for the study of Human Evolution. 10-12 September. 2015. http://www.eshe.eu/static/eshe/files/PESHE/PESHE_4_2015_London.pdf
David Reich, Richard E. Green, Martin Kircher, Johannes Krause, Nick Patterson, Eric Y. Durand, Bence Viola, Adrian W. Briggs, Udo Stenzel, Philip L. F. Johnson, Tomislav Maricic, Jeffrey M. Good, Tomas Marques-Bonet, Can Alkan, Qiaomei Fu, Swapan Mallick, Heng Li, Matthias Meyer, Evan E. Eichler, Mark Stoneking, Michael Richards, Sahra Talamo, Michael V. Shunkov, Anatoli P. Derevianko, Jean-Jacques Hublin, Janet Kelso, Montgomery Slatkin, Svante Pääbo. Genetic history of an archaic hominin group from Denisova Cave in Siberia // Nature. 2010. V. 468. P. 1053–1060.
Amar J. Majmundar, Waihay J. Wong & M. Celeste Simon (October 2010). Hypoxia-inducible factors and the response to hypoxic stress// Molecular cell 40 (2): 294–309.
Shuhua Xu, Shilin Li, Yajun Yang, Jingze Tan, Haiyi Lou, Wenfei Jin, Ling Yang, Xuedong Pan, Jiucun Wang, Yiping Shen, Bailin Wu, Hongyan Wang, and Li Jin A Genome-Wide Search for Signals of High-Altitude Adaptation in Tibetans Mol Biol Evol (2011) 28(2): 1003—1011 first published online October 20, 2010
Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S, Schraiber JG, Jay F, Prüfer K, de Filippo C, Sudmant PH, Alkan C, Fu Q, Do R, Rohland N, Tandon A, Siebauer M, Green RE, Bryc K, Briggs AW, Stenzel U, Dabney J, Shendure J, Kitzman J, Hammer MF, Shunkov MV, Derevianko AP, Patterson N, Andrés AM, Eichler EE, Slatkin M, Reich D, Kelso J, Pääbo S. A high-coverage genome sequence from an archaic Denisovan individual// Science. 2012 Oct 12;338(6104):222-6. Epub 2012 Aug 30. http://www.ncbi.nlm.nih.gov/pubmed/22936568?dopt=Abstract&holding=npg
Haak W., Lazaridis I., Patterson N., Rohland N., Mallick S., Llamas B., Brandt G., Nordenfelt S., Harney E., Stewardson K., Fu Q., Mittnik A., Bánffy E., Economou C., Francken M., Friederich S., Garrido Pena R., Hallgren F., Khartanovich V., Khokhlov A., Kunst M., Kuznetsov P., Meller H., Mochalov O., Moiseyev V., Nicklisch N., Pichler S., Risch R., Rojo Guerra M.,Roth C., Szécsényi-Nagy A., Wahl J., Meyer M., Krause J., Brown D., Anthony D., Cooper A., Werner Alt K., Reich D. Massive migration from the steppe is a source for Indo-European languages in Europe//Nature. 2015 June 11; 522(7555): 207-11
Allentoft M., Sikora M., Sjögren K., Rasmussen S., Rasmussen M., Stenderup J., Damgaard P., Schroeder H.,Ahlström T.,Vinner L., Malaspinas A., Margaryan A., Higham T., Chivall D., Lynnerup N., Harvig L., Baron J., Della Casa P., Dąbrowski P., Duffy P., Ebel A., Epimakhov A., Frei K., Furmanek M., Gralak T., Gromov A., Gronkiewicz S., Grupe G., Hajdu T., Jarysz R., Khartanovich V., Khokhlov A., Kiss V., Kolář J., Kriiska A., Lasak I., Longhi C., McGlynn G., Merkevicius A., Merkyte I., Metspalu M., Mkrtchyan R., Moiseyev V., Paja L., Pálfi G., Pokutta D., Pospieszny L., Douglas Price T., Saag L., Sablin M., Shishlina N., Smrčka V., Soenov V., Szeverényi V., Tóth G., Trifanova S., Varul L., Vicze M., Yepiskoposyan L., Zhitenev V., Orlando L., Sicheritz-Pontén T., Brunak S., Nielsen R., Kristiansen K., Willerslev E. Population genomics of Bronze Age Eurasia//Nature 522, 167–172 (11 June 2015).
Yu-Sheng Chen, Antonel Olckers, Theodore G. Schurr, Andreas M. Kogelnik, Kirsi Huoponen, and Douglas C. Wallace. mtDNA Variation in the South African Kung and Khwe—and Their Genetic Relationships to Other African Populations. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1288201/ http://www.ianlogan.co.uk/discussion/hap_G.htm http://www.ianlogan.co.uk/discussion/hap_C.htm
Clio Der Sarkissian, Paul Brotherton, Oleg Balanovsky, Jennifer E. L. Templeton, Bastien Llamas, Julien Soubrier, Vyacheslav Moiseyev, Valery Khartanovich, Alan Cooper, Wolfgang Haak, The Genographic Consortium Mitochondrial Genome Sequencing in Mesolithic North East Europe Unearths a New Sub-Clade within the Broadly Distributed Human Haplogroup C1//Plosone Published: February 4, 2014DOI: 10.1371/journal.pone.0087612 http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0087612
Alexey G. Nikitin, Jeremy R. Newton, Inna D. Potekhina Mitochondrial haplogroup C in ancient mitochondrial DNA from Ukraine extends the presence of East Eurasian genetic lineages in Neolithic Central and Eastern Europe//Journal of Human Genetics 57, 610-612 (September 2012) | doi:10.1038/jhg.2012.69. http://www.nature.com/jhg/journal/v57/n9/full/jhg201269a.html
David Comas, Stephanie Plaza, R. Spencer Wells, Nadira Yuldaseva, Oscar Lao, Francesc Calafell, Jaume Bertranpetit Admixture, migrations, and dispersals in Central Asia: evidence from maternal DNA lineages//European Journal of Human Genetics (2004) 12, 495–504. doi:10.1038/sj.ejhg.5201160. Published online 11 February 2004. http://www.nature.com/ejhg/journal/v12/n6/pdf/5201160a.pdf
David Comas, Stephanie Plaza, R. Spencer Wells, Nadira Yuldaseva, Oscar Lao, Francesc Calafell, Jaume Bertranpetit Admixture, migrations, and dispersals in Central Asia: evidence from maternal DNA lineages//European Journal of Human Genetics (2004) 12, 495–504. doi:10.1038/sj.ejhg.5201160. Published online 11 February 2004. http://www.nature.com/ejhg/journal/v12/n6/pdf/5201160a.pdf
Amory S., Crubézy E., Keyser C., Alekseev A.N., Ludes B. Early influence of the steppe tribes in the peopling of Siberia//Human Biology. October 2006. 78 (5): 531–49. Aleksandr S. Pilipenko, Rostislav O. Trapezov, Anton A. Zhuravlev, Vyacheslav I. Molodin, Aida G. Romaschenko. MtDNA Haplogroup A10 Lineages in Bronze Age Samples Suggest That Ancient Autochthonous Human Groups Contributed to the Specificity of the Indigenous West Siberian Population//Plosone. Published: May 7, 2015DOI: 10.1371/journal.pone.0127182 http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0127182 http://www.ianlogan.co.uk/discussion/hap_F.htm Ruiz-Pesini E, Lapeña AC, Díez-Sánchez C, et al. Human mtDNA haplogroups associated with high or reduced spermatozoa motility. American Journal of Human Genetics. 67 (3): 682–96. September 2000. DOI:10.1086/303040. PMID 10936107.
Achilli et al., The Molecular Dissection of mtDNA Haplogroup H Confirms That the Franco-Cantabrian Glacial Refuge Was a Major Source for the European Gene Pool//American Journal of Human Genetics, 2004 November; 75(5): 911.
Березина Г.М., Святова Г.С., Абдуллаева А.М. Бермишева М.А., Кутуев И.А., Хуснутдинова Э.К., Виллемс Р. Полиморфизм митохондриальной ДНК в казахской популяции//Медицинская генетика. 2005. Том 4. № 3. С. 108-113. https://sites.google.com/site/pinhasierc/home/samples
Keyser C., Bouakaze C., Crubézy E., Nikolaev V., Montagnon D., Reis T., Ludes B. Ancient DNA provides new insights into the history of south Siberian Kurgan people//Human Genetics (2009). 126:395–410. http://www.snpedia.com/index.php/Rs12913832
Allentoft M., Sikora M., Sjögren K., Rasmussen S., Rasmussen M., Stenderup J., Damgaard P., Schroeder H.,Ahlström T.,Vinner L., Malaspinas A., Margaryan A., Higham T., Chivall D., Lynnerup N., Harvig L., Baron J., Della Casa P., Dąbrowski P., Duffy P., Ebel A., Epimakhov A., Frei K., Furmanek M., Gralak T., Gromov A., Gronkiewicz S., Grupe G., Hajdu T., Jarysz R., Khartanovich V., Khokhlov A., Kiss V., Kolář J., Kriiska A., Lasak I., Longhi C., McGlynn G., Merkevicius A., Merkyte I., Metspalu M., Mkrtchyan R., Moiseyev V., Paja L., Pálfi G., Pokutta D., Pospieszny L., Douglas Price T., Saag L., Sablin M., Shishlina N., Smrčka V., Soenov V., Szeverényi V., Tóth G., Trifanova S., Varul L., Vicze M., Yepiskoposyan L., Zhitenev V., Orlando L., Sicheritz-Pontén T., Brunak S., Nielsen R., Kristiansen K., Willerslev E. Population genomics of Bronze Age Eurasia//Nature 522, 167–172 (11 June 2015).
База данных современных полных сиквенсов Y-хромосомы гаплогруппы R1a. http://www.yfull.com/tree/R1a/
Mathieson I., Lazaridis I., Rohland N., Mallick S., Patterson N., Roodenberg S.A., Harney E., Stewardson K., Fernandes D., Novak M., Sirak K., Gamba C., Jones E.R., Llamas B., Dryomov S., Pickrell J., Arsuaga J.L., Bermúdez de Castro J.M., Carbonell E., Gerritsen F., Khokhlov A., Kuznetsov P., Lozano M., Meller H., Mochalov O., Moiseyev V., Rojo Guerra M.A., Roodenberg J., Vergès J.M., Krause J., Cooper A., Alt K.W., Brown D., Anthony D., Lalueza-Fox C., Haak W., Pinhasi R., Reich D. Eight thousand years of natural selection in Europe. Available at: http://www.biorxiv.org/content/biorxiv/early/2015/10/10/016477.full.pdf https://docs.google.com/spreadsheets/d/1Vjbp450AwI7R-Y9J1YGSm9FjJWu9s9lx1azjUJbS8hQ/edit?pli=1#gid=953757259
Haak W., Lazaridis I., Patterson N., Rohland N., Mallick S., Llamas B., Brandt G., Nordenfelt S., Harney E., Stewardson K., Fu Q., Mittnik A., Bánffy E., Economou C., Francken M., Friederich S., Garrido Pena R., Hallgren F., Khartanovich V., Khokhlov A., Kunst M., Kuznetsov P., Meller H., Mochalov O., Moiseyev V., Nicklisch N., Pichler S., Risch R., Rojo Guerra M.,Roth C., Szécsényi-Nagy A., Wahl J., Meyer M., Krause J., Brown D., Anthony D., Cooper A., Werner Alt K., Reich D. Massive migration from the steppe is a source for Indo-European languages in Europe//Nature. 2015 June 11; 522(7555): 207-11
Mathieson I., Lazaridis I., Rohland N., Mallick S., Patterson N., Roodenberg S.A., Harney E., Stewardson K., Fernandes D., Novak M., Sirak K., Gamba C., Jones E.R., Llamas B., Dryomov S., Pickrell J., Arsuaga J.L., Bermúdez de Castro J.M., Carbonell E., Gerritsen F., Khokhlov A., Kuznetsov P., Lozano M., Meller H., Mochalov O., Moiseyev V., Rojo Guerra M.A., Roodenberg J., Vergès J.M., Krause J., Cooper A., Alt K.W., Brown D., Anthony D., Lalueza-Fox C., Haak W., Pinhasi R., Reich D. Eight thousand years of natural selection in Europe. Available at: http://www.biorxiv.org/content/biorxiv/early/2015/10/10/016477.full.pdf https://docs.google.com/spreadsheets/d/1Vjbp450AwI7R-Y9J1YGSm9FjJWu9s9lx1azjUJbS8hQ/edit?pli=1#gid=953757259
Mathieson I., Lazaridis I., Rohland N., Mallick S., Patterson N., Roodenberg S.A., Harney E., Stewardson K., Fernandes D., Novak M., Sirak K., Gamba C., Jones E.R., Llamas B., Dryomov S., Pickrell J., Arsuaga J.L., Bermúdez de Castro J.M., Carbonell E., Gerritsen F., Khokhlov A., Kuznetsov P., Lozano M., Meller H., Mochalov O., Moiseyev V., Rojo Guerra M.A., Roodenberg J., Vergès J.M., Krause J., Cooper A., Alt K.W., Brown D., Anthony D., Lalueza-Fox C., Haak W., Pinhasi R., Reich D. Eight thousand years of natural selection in Europe. Available at: http://www.biorxiv.org/content/biorxiv/early/2015/10/10/016477.full.pdf https://docs.google.com/spreadsheets/d/1Vjbp450AwI7R-Y9J1YGSm9FjJWu9s9lx1azjUJbS8hQ/edit?pli=1#gid=953757259
Hollard C., Keyser C., Giscard P.-H., Tsagaan T., Bayarkhuu N., Bemmann J., Crubezy E., Ludes B. Strong genetic admixture in the Altai at the Middle Bronze Age revealed by uniparental and ancestry informative markers//Forensic Science International: Genetics, 2014, № 12, pp. 199-207.
Zhao Y.-B., Zhang Y., Li H.-J., Cui Y.-Q., Zhu H., Zhou H. Ancient DNA evidence reveals that the Y chromosome haplogroup Q1a1 admixed into the Han Chinese 3,000 years ago// American Journal of Human Biology. November/December 2014, Volume 26, Issue 6, pp. 813–821. http://iggc.kz/wp-content/uploads/2017/03/Rezultaty-raboty-Lab-Pop-Gen-noyab-2016.pdf http://www.yfull.com/tree/Q/ http://www.isogg.org/tree/ISOGG_HapgrpC.html
